
Current environmental changes are often considered as negatively impacting the affective state of animals.
Yet, the interplay betweenenvironmental conditions and affective state should rather be viewed as a reciprocal and dynamic relationship, as variation in affective state likely determines how animals decide to respond to environmental changes.
Here, I illustrate how affective states may contribute to determining how animals respond to environmental changes through phenotypic plasticity, environmental modification or dispersal. A condition for future studies to examine this hypothesis will be to consider the response of animals in parallel with valid indicators of affective state reflecting different affective dimensions (valence, arousal) over different timescales.
Moreover, considering affective state as a central component of the response of animals to environmental changes implies that a condition to realistically investigate this response is to provide animals with the freedom to decide between options associated with different affective values.
The role of affective state in decision-making
(a) Decision-making: from non-sentient to sentient organisms
(b) The affective model of decision-making
(c) The affective model of decision-making and the response of animals to environmental changes
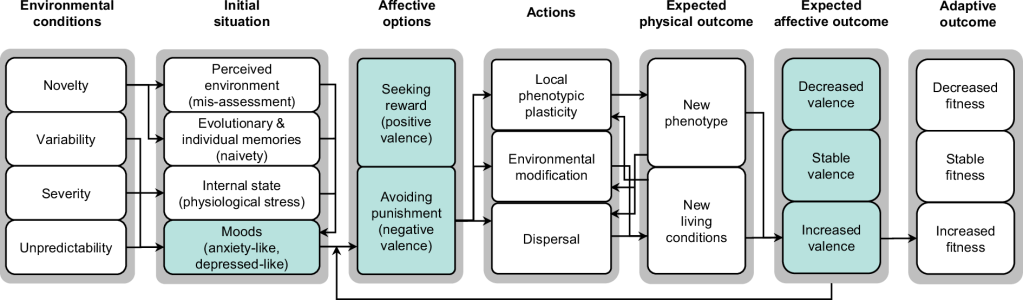
Green elements are specific to sentient animals, while white elements are also present in non-sentient organisms.
The arrows between elements have an illustrative purpose and represent a possible pathway explaining how decisions are made in sentient animals in response to the general characteristics of current environmental changes (other pathways are possible). Terms in parentheses describe the expected initial situation of animals responding to challenging environmental changes.
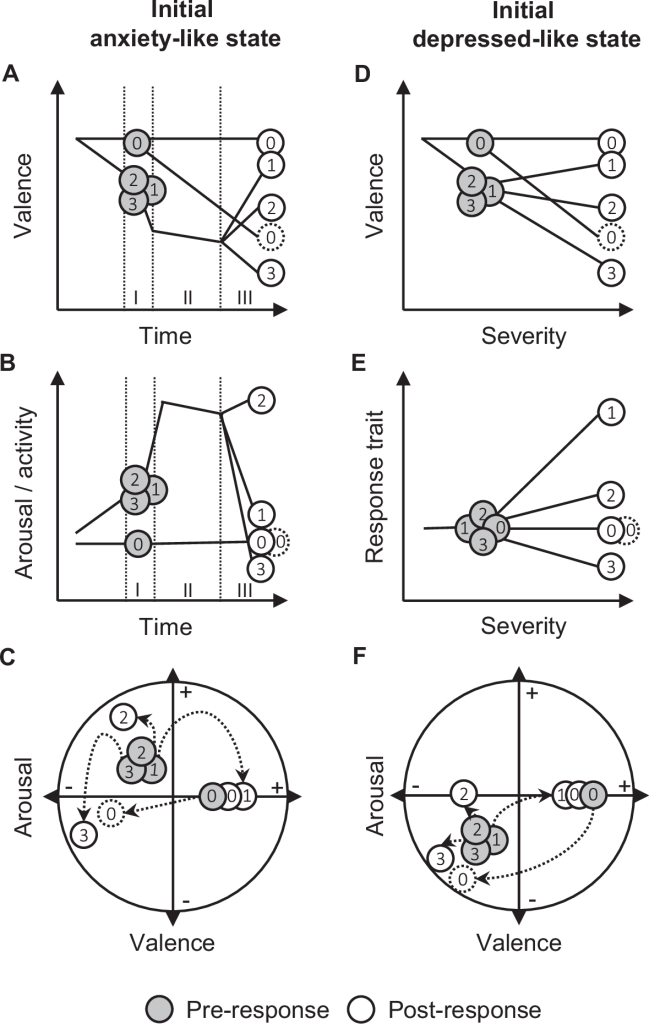
The resulting negatively valenced affective states individuals initially experience are either accompanied by an increased arousal (anxiety-like state; A–C) or a decreased arousal (depressed-like state; D–F). For the responses following an initial anxiety-like state, valence and arousal (movements) are represented as a function of time to visualize the successive phases of the response (e.g. dispersal: departure (I), transience (II), settlement (III);
A, B). For the responses following an initial depressed-like state, valence and expression of response trait are represented as a function of the severity of environmental conditions (using a reaction norm representation;
D, E). In both cases, variation in valence and arousal are also represented in affective space (C, F).
Grey symbols and white symbols, respectively, represent individuals before and after responding to environmental changes. Individual 0 (control) does not respond due to the non-necessity to respond (stable valence; white symbol with solid line) or its inability to do so (decreasing valence; white symbol with dashed line).
Individuals 1–3 respond to different degrees, resulting in different mitigating effects on their valence (individual 1: complete mitigation; individual 2: partial mitigation; individual 3: no mitigation).
Practical considerations and perspectives
(a) Considering affective state and response together
The affective state experienced by wild animals has not been considered so far to explain how they respond to current environmental changes.
The aim of this article is to start filling this gap by stimulating interest in the role of affective state in shaping the response of animals to environmental changes and by encouraging future studies to examine this point.
A key aspect for future studies to empirically examine how the affective state of animals interacts with their response to environmental changes will be to monitor how it varies over the successive phases of a local (i.e. before, during, after) or dispersing response (i.e. departure, transience, settlement), and how it differs between individuals adopting different responses and responding to different degrees.
This temporal monitoring could be conducted for valence and arousal separately (figure 2A, B) but also more comprehensively by using affective space representations (figure 2C, F). In addition, to identify the traits involved in the response of animals and that interact with their affective state, it would be useful to consider affective states in parallel with these response traits in terms of reaction norms (i.e. the range of valence values and response trait values expressed by animals across variable conditions; figure 2D, E). In this line of thought, it would also be interesting to examine how affective states covary and interact with the set of morphological, physiological and behavioural traits already known to correlate with the propensity of animals to disperse.
(b) Using proper indicators of affective state
Examining the response of animals to environmental changes using the affective model of decision-making inherently requires the use of an affective framework where affective states represent the core of animal welfare and affective valence its fundamental currency.
In contrast to other theoretical frameworks previously used to assess animal welfare (e.g. naturalness of living conditions, biological functioning), adopting an affective perspective strongly complicates welfare assessment because of the inherent inaccessibility of privately experienced affective states.
However, affective states do not occur in isolation but depend on different physical and functional factors (environment, nutrition, health, behaviour) and, in turn, can affect a variety of physiological, behavioural and cognitive markers that could be used as indicators of affective states. These potential indicators differ in a variety of properties, including their ability to reflect valence and arousal over different timescales.
These differences are important to consider when examining the response of animals to current environmental changes, as the decisions made by animals are expected to be based on both long-term negatively valenced
affective states characterized with different levels of arousal (anxiety-like and depressed-like states), and short-term affective states that vary both in terms of valence and arousal (e.g. excitement-like, relief-like, contentment-like, nervousness-like, frustration-like states).
Examining the response of animals to environmental changes based on their affective state, therefore, requires using both long-term indicators of affective state reflecting the cumulative experience of animals before the decision is made (e.g. telomere length, attention bias) and more plastic ones reflecting affective changes during and following decisionmaking (e.g. oxidative status, postures, vocalizations).
These markers should also cover both negative and positive valences as well as low and high levels of arousal (figure 2). Most importantly, before being used, these markers need to be validated for their ability to reflect affective states. Towards this end, examining how markers vary across multiple conditions predicted to be associated with the same valence and across multiple conditions predicted to be associated with the opposite valence may be useful.
For instance, finding that markers vary in a consistent way across negatively valenced conditions while consistently varying in the opposite direction across positively valenced conditions suggests that these markers can be used as valid indicators of affective state in the considered species irrespective of the specific conditions altering the affective state of animals.
(c) Giving animals the freedom to decide
A logical consequence of considering affective state as a central component of the response of animals to environmental changes is that animals should have the opportunity to choose between options predicted to have different affective values when examining this response.
This objective might be difficult to attain when using experimental approaches in captivity where animals are typically forced to experience specific conditions locally with not much behavioural flexibility to decide on (potentially leading to a depressed-like state).
Studies conducted in the wild, therefore, appear necessary to at least complement captive studies, as they offer animals full freedom on how to decide between all possible options and, therefore, provide a more realistic view of how the affective state of animals interacts with their response to environmental changes.
