
From competition to facilitation and mutualism: a general theory of the niche by Koffel, Daufresne and Klausmeier explores the niche theory.
Niche Theory is a central framework in ecology based on the recognition that most interactions between organisms are indirect, mediated by the biotic and abiotic dynamical environment these organisms live in. Despite its potential generality, the theory still mostly focuses on how resource–consumer dynamics mediate competition in ecological communities.
However, it is being increasingly recognized that positive interactions between organisms also play an important role in driving the structure and functioning of ecological communities, from plants to microbes. A unified theory of the niche is presented, that applies to both positive and negative interactions between organisms, mediated by one or two environmental factors.Classical concepts such as niche differences and fundamental and realized niches can naturally be expanded to facilitative and mutualistic interactions.
New general niche concepts that appear exclusively in the presence of positive interactions are:
(1) the Allee niche, a region of environmental conditions for which a species can persist but not invade from low densities and
(2) niche facilitation, when the presence of a species expands the set of environmental conditions under which a second species can invade and/or persist.
The broad applicability of this theory is illustrated by these concepts using a diverse set of theoretical examples, from bacteria feeding on an inhibiting substrate, to nitrogen-fixing plants and the indirect mutualism between a plant and a carnivore species.
In sum, Niche Theory provides a natural framework for positive interactions in ecology, bringing a unified perspective and new conceptual tools to study ecological systems where these positive interactions occur.
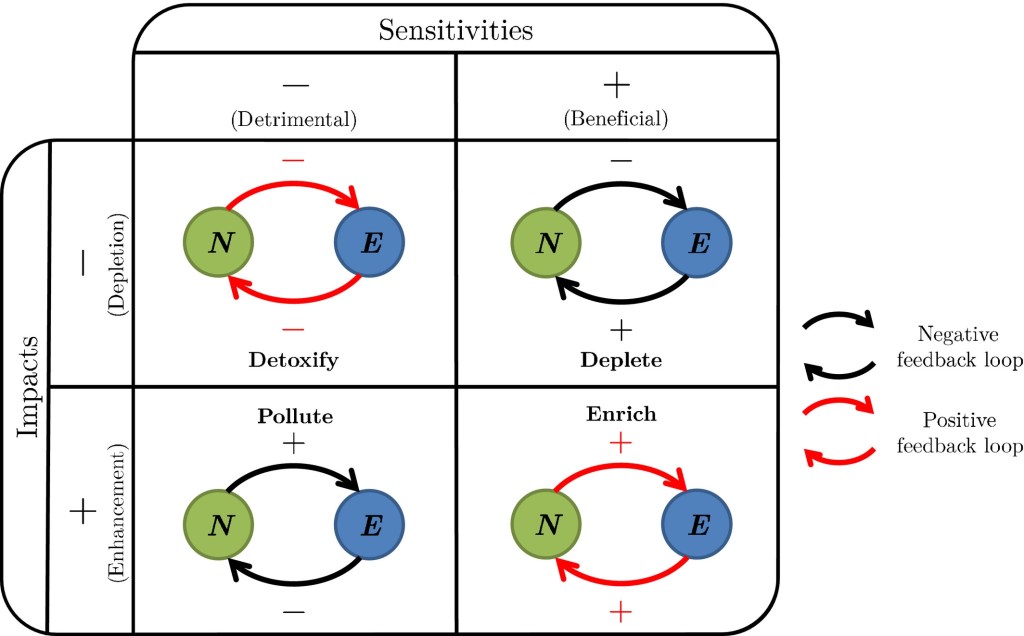
in “Deplete,” focal organisms are bottom-up controlled as they consume (−impact) a resource needed for growth (+sensitiv) and, in “Pollute,” they are top-down controlled as they feed (+impact) an enemy that hurts them (−sensitiv).
The two other scenarios lead to positive feedback loops, i.e., unregulated growth (red): in “Enrich,” organisms produce (+impact) a resource they need to grow (+sensitivity) and in “Detoxify,” organisms deplete (−impact) a resource that inhibits their growth (−sensitivity).
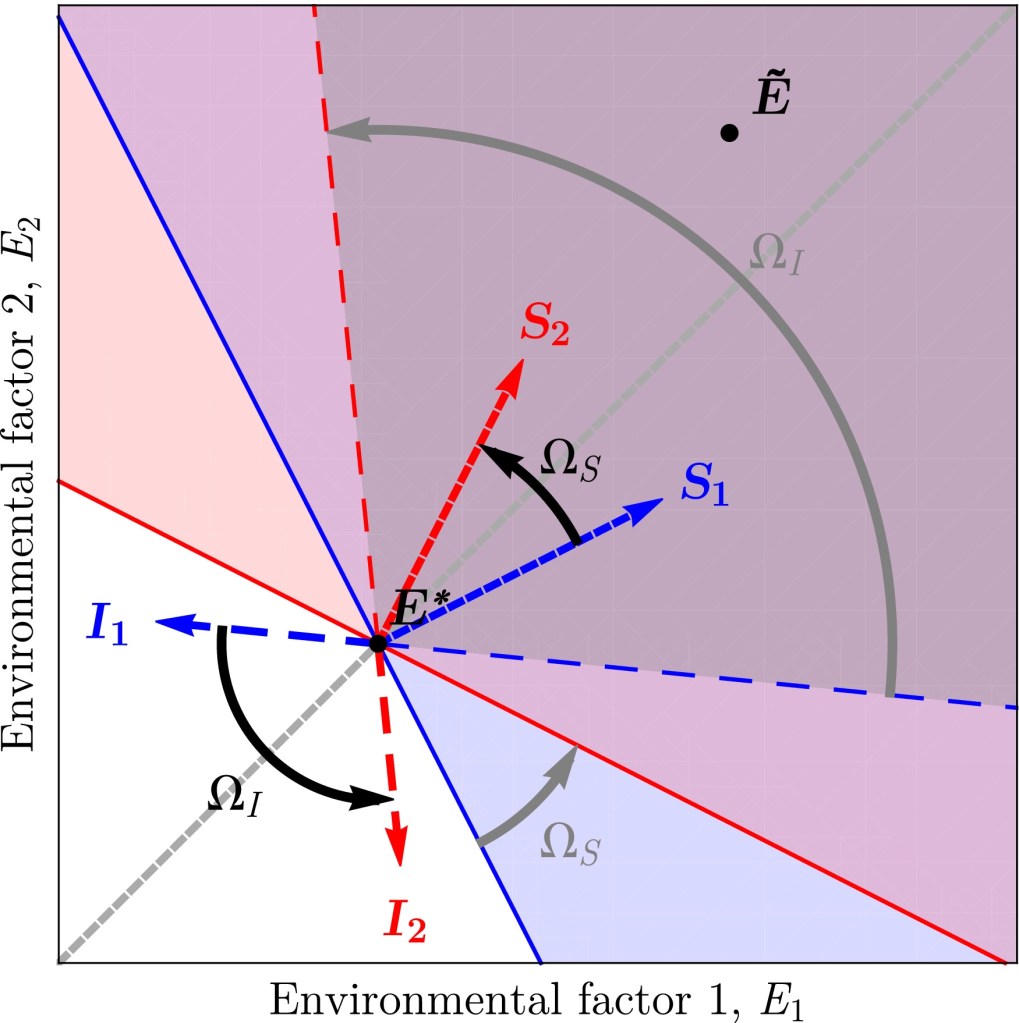


(A) Setup more general than the symmetrical case, as the centroid of the coexistence region (gray, dashed) is tilted relatively to the centroid of the sensitivity vectors (solid, brown) by an angle θ. As a consequence, interactions between the two species are overall less detrimental/more beneficial to one of the two species (on this particular example, species 2 is favored over 1).
(B) As a consequence of this asymmetry, intermediate sensitivity and impact niche differences lead to a new regime of “(−, +) coexistence” where species 1 facilitates species 2 while species 2 competes with species 1 (θ = 11.25° in this particular example).
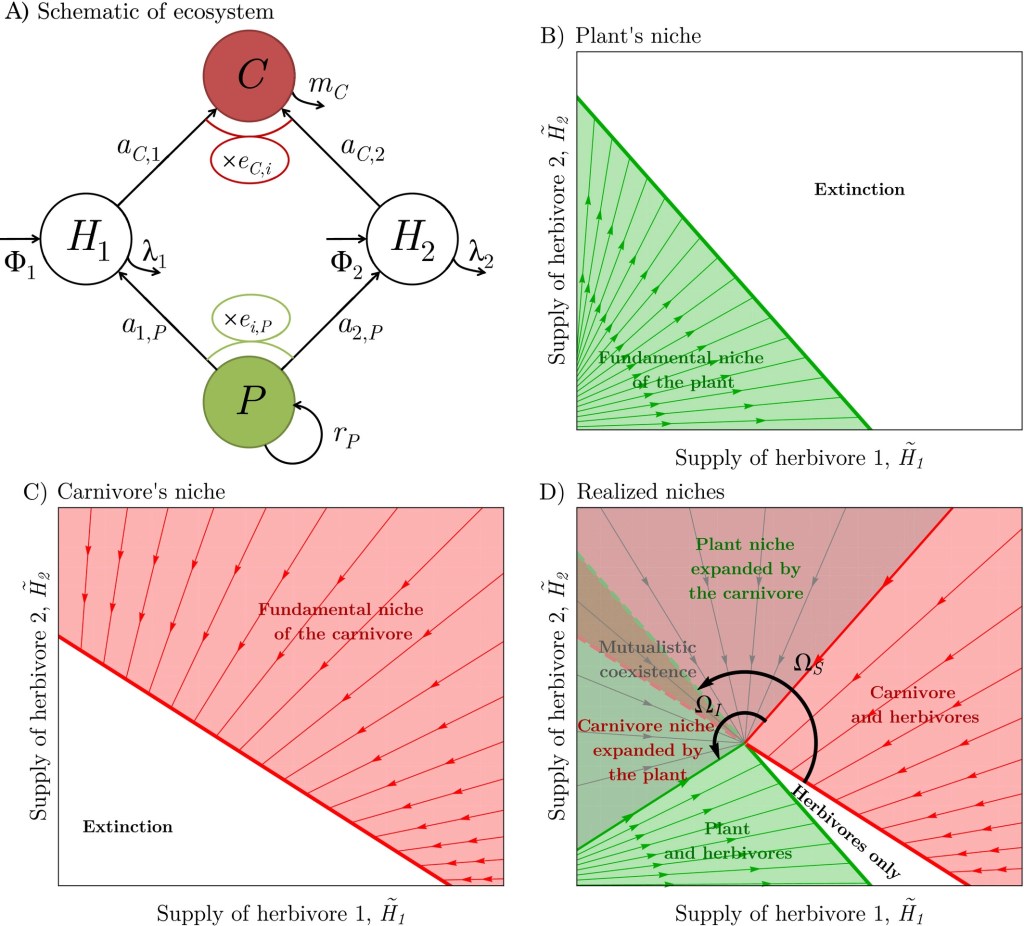
(A) The plant (P, green) benefits the carnivore (C, red) by feeding the herbivores, and the carnivore benefits the plant by consuming the herbivores. Local herbivore density does not rely solely on the focal plant, as it is partially controlled by immigration from the surrounding landscape, through the rates Φ1 and Φ2. Herbivores differ in their relative “quality”: herbivore 1 strongly grazes the focal plant and does not feed the carnivore well, while herbivore 2 damages the plant less and feeds the carnivore better.
(B) fundamental niche of the plant along gradients of herbivore immigration. The plant can only persist below a certain level of total herbivore immigration, materialized by its linear ZNGI (green, thick line).
(C) Fundamental niche of the carnivore. The carnivore can only persist above a certain level of total herbivore immigration, materialized by its linear ZNGI (red, thick line).
(D) Realized niches of the plant and the carnivore in interaction. Under low herbivore immigration, the plant can persist as the carnivore is excluded (green region). Similarly, high herbivore immigration enables the carnivore to persist in the absence of the plant (red region). Under intermediate immigration regimes that favor the relative abundance of herbivore 1, both the plant and the carnivore go locally extinct because herbivores are too abundant from the plant perspective, but not abundant enough from the carnivore perspective (white region). Conversely, under immigration that favors the relative abundance of herbivore 2, the plant and carnivore coexist together alongside the herbivores (gray region), strongly facilitating each other by expanding their niches. There, plant and carnivore biomass reach high levels, as indicated by an impact niche difference close to 180° on this example.
This paper advocates for the development of a general theory of species interactions. More particularly, the framework of Niche Theory specifically tailored to indirect interactions mediated by explicit environmental factors, shows how a concept like niche difference, usually used to quantify the strength of competition, can equally be applied to situations that involve positive interspecific interactions. This approach suggests building on the connection between lower-level metrics of direct interactions between the focal species and their environment (the sensitivity and impact niche differences) and an effective, indirect metric of niche difference. With several possible options to pick from for the latter, we identified pros and cons for each metric are identified, which suggests that there is no ideal metric to measure effective niche difference.
