
The feature review in TICS “Evolutionary and neuroendocrine foundations of human aggression” by Amar Sarkar, and Richard Wrangham describes a human behavioural paradox: they are peaceful in many circumstances, but they are also violent and kill conspecifics at high rates.
The review describes a social evolutionary theory to resolve this paradox. The theory interprets human aggression as a combination of low propensities for reactive aggression and coercive behaviour and high propensities for some forms of proactive aggression (especially coalitionary proactive aggression).
These tendencies are associated with the evolution of groupishness, self-domestication, and social norms.
This human aggression profile is expected to demand substantial plasticity in the evolved biological mechanisms responsible for aggression. The review discusses the contributions of various social signalling molecules (testosterone, cortisol, oxytocin, vasopressin, serotonin, and dopamine) as the neuroendocrine foundation conferring such plasticity.
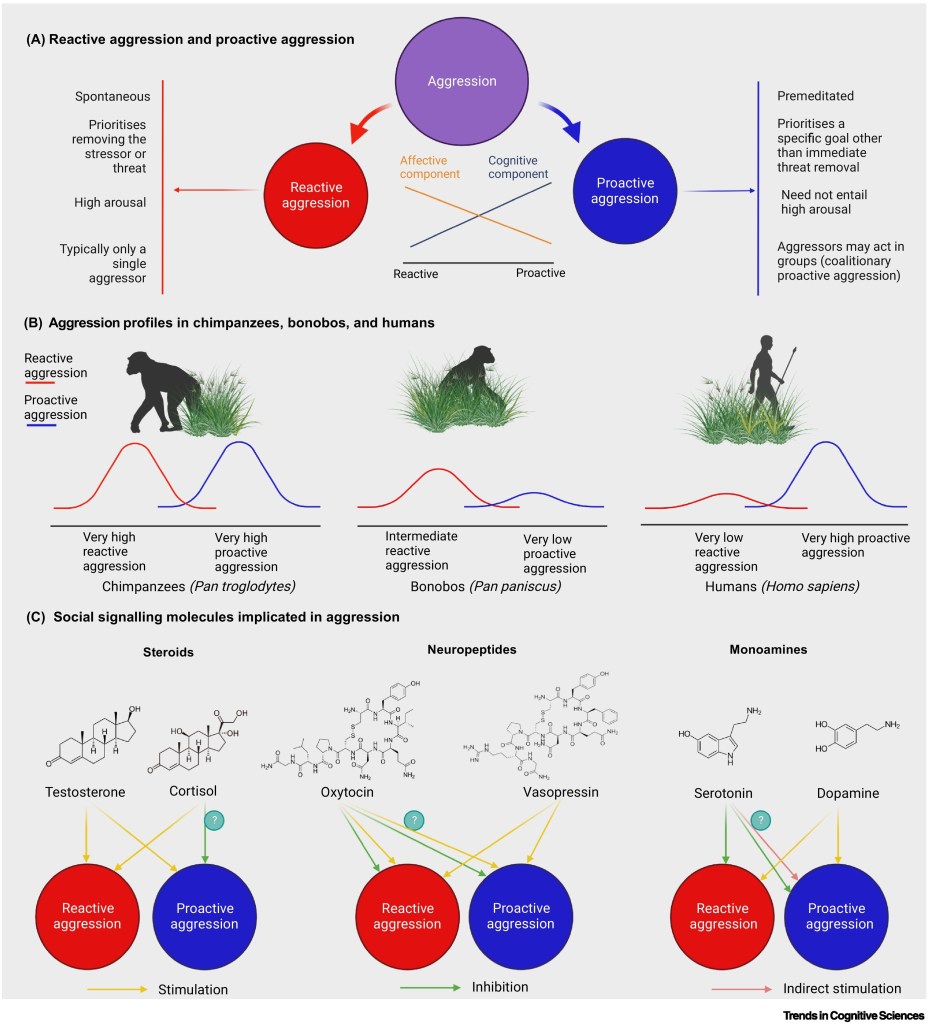
(A) Features of reactive and proactive aggression.
(B) Hominids differ dramatically in species-typical tendencies for reactive and proactive aggression. Uncertainty arises over how the frequency and nature of reactive aggression in bonobos compare to those of chimpanzees. Male bonobos are clearly less violent than chimpanzees in interactions with females and in intergroup encounters. However, female bonobos readily resort to aggression against competing or bullying males, whilst unpublished data suggest that male bonobos have high rates of intragroup aggression against males, most of which is probably reactive.
(C) Social signalling molecules are associated with reactive and proactive aggression. We illustrate how each social signalling molecule may interact with aggression. For example, cortisol appears to be positively related to reactive aggression (at least in the short term), but is either unrelated or negatively related to proactive aggression (we depict a negative association, but indicate the uncertainty with the question mark). Oxytocin is, on average, negatively associated with reactive aggression, but is positively associated with reactive aggression in infant defence. Oxytocin appears to be negatively associated with many forms of proactive aggression, but positively associated with coalitionary proactive aggression. However, it is unclear whether this positive correlation is a direct association or an indirect association with the various social aspects of coalitionary aggression (we depict a positive association, but indicate the uncertainty with the question mark). Serotonin is negatively associated with reactive aggression and some forms of proactive aggression, but is positively associated with executive functions that could support coalitionary proactive aggression (we depict an indirect association, but indicate the uncertainty with the question mark). For simplicity, we depict causal molecule→behaviour associations, though the evidence is often correlational rather than experimental.
Self-domestication
Self-domestication is proposed to have occurred in several species, including humans and bonobos. Evidence for self-domestication arises from parallels with domestication. Researchers since at least Darwin have observed characteristic anatomical, physiological, and behavioural differences between domesticated mammals and their wild progenitors and relatives. Such features (~30 traits) constitute the domestication syndrome. For instance, domesticates commonly display shorter faces, smaller molars, curled tails, more white fur, smaller brains, reduced weaponry, attenuated corticosteroid responses, paedomorphism, or increased play. However, such features can occur unpredictably. For example, although domesticates tend to have more white fur patches than their wild relatives, some individuals within the species do not have any white fur patches (e.g., dogs). Likewise, some domesticated species lack any white fur (e.g., water buffalo). Such trends have led some researchers to question the existence of the domestication syndrome. However, given that such traits recur across domesticates (and none of these traits was the target of selection), the domestication syndrome can be regarded as a robust phenomenon.
Controlled breeding of foxes, mink, rodents, and chickens points to the pattern that selection against reactive aggression drives domestication syndromes. Experiments targeting low reactive aggression reveal a principal cause of domestication syndrome: selection for individuals responding to potential threats with unusually low fear and aggression. Notably, within a few generations of selection for reduced aggression (the target of selection), captive foxes in Russia showed increased frequencies of white fur patches, curled tails, and other features of a domestication syndrome. Each of these traits can be interpreted as a correlated byproduct, a spandrel, rather than the target of selection.
Inferring histories of domestication and selection against reactive aggression
Controlled breeding experiments demonstrate that selection against reactive aggression can elicit features of a domestication syndrome. Importantly, domestication and self-domestication should both elicit a domestication syndrome. In other words, it does not appear to matter whether such selection occurs through human intervention (e.g., cattle) or over natural evolution (e.g., bonobos): we can expect features of a domestication syndrome whenever species experience selection against reactive aggression, such that descendants are less aggressive than their ancestors. This insight proves useful for reconstructing evolutionary events: given evidence of a domestication syndrome, selection probably acted against reactive aggression in the history of that species. Interestingly, selecting for fighting ability in female cattle (performed to produce animals for contests and shows) appears to dampen or reverse some aspects of domestication, including reducing social tolerance, increasing propensity to fight spontaneously with conspecifics, and increasing endogenous testosterone concentrations.
Anatomical changes in H. sapiens from its immediate Pleistocene ancestor (often referred to as H. heidelbergensis) strongly suggest that features of domestication syndrome had emerged by ~300 000 years ago. Since humans display strikingly low reactive aggression, we infer that selection against reactive aggression (e.g., via targeted conspiratorial killing of coercive alpha males) drove human self-domestication.
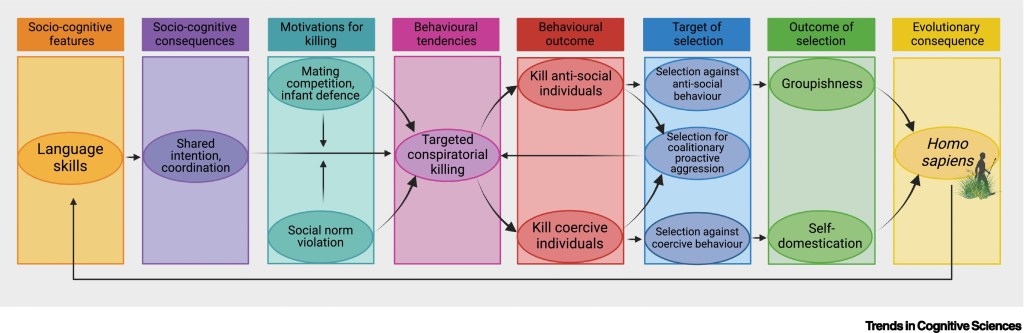
Targeted conspiratorial killing is hypothesised to have driven the emergence of both selfdomestication and groupishness, creating the anatomical and psychological background for the evolution of Homo sapiens.
The motivations driving targeted conspiratorial killing could be purely fitness-oriented without reference to social norms, such as increasing access to mates or preventing infanticide by killing coercive alpha males. Alternatively, they could be more norm-oriented, and entail punishment of individuals who persistently or egregiously violate social norms (such as coercive alpha males). In the upper pathway, targeted conspiratorial killing leads to the social control of antisocial individuals, strong selection against antisocial behaviour, strong selection for costly prosocial behaviour, and the emergence of groupishness.
In the lower pathway, targeted conspiratorial killing leads to the social control of coercive individuals (selecting against reactive aggression and some forms of proactive aggression), and the emergence of self-domestication. In both cases, alliance-mediated killing of antisocial and coercive individuals also maintains selection for coalitionary proactive aggression.
Development of a uniquely sophisticated form of vocal signalling (language) is hypothesised to generate a highly sensitive and specific channel of communication that enables fine-grained sharing of intentions and behavioural coordination.
The same linguistic ability also enables fine-grained communication about the extent of social norm violation and reproductive control exerted by coercive males.
Groupishness
Compared to other mammals (including primates), the extraordinary nature and scale of human cooperation are thought to be unique. However, it has been difficult to specify precisely how humans differ from other species in this respect. Much of human exceptionalism regarding cooperation derives from H. sapiens being the only species in which individuals routinely assist others without predictable payoffs from kin selection, mutualism, or direct reciprocity. In their absence, the only mechanism by which the agent’s aid can be repaid is if a third party returns a benefit. A special feature of human cooperation is thus a tendency to act prosocially in ways that are adaptive only because of subsequent third-party involvement. This tendency is not typically defined with any particular term, and we refer to it as ‘groupishness’.
Cooperation that is favoured only because of third-party involvement (groupishness, in our usage) has often been thought of as having evolved in a system of indirect reciprocity. Indirect reciprocity theory suggests that a generous agent benefits from later rewards because the agent has earned a positive reputation for helping. Third parties, aware of the agent’s positive reputation, are willing to pay costs to reward the agent’s earlier prosocial acts. One difficulty with this theory is that evidence of indirect reciprocity occurring in naturalistic contexts (as opposed to laboratory experiments) is currently limited.
More recently, signalling theory has been argued to explain such cooperation better than (or alongside) indirect reciprocity, partly because it shows that third parties can benefit by rewarding the actor without paying costs to do so. Applying signalling theory to the phenomenon of groupishness, the agent’s prosocial behaviour signals their cooperative quality as a norm-follower. Signals of the agent’s quality lead third parties to choose to interact positively with the agent, which the third parties do for their own selfish reasons. In support of signalling theory, individuals have often been shown to manipulate their reputations in ways that signal their quality. For example, children are more prosocial in public than in private settings by 5 years of age.
Signalling theory readily supplies explanations for the evolution of groupishness: when alpha alliances seized control of their social groups, they created or strengthened norms that benefitted their interests and punished norm violators. Under such circumstances, displays of conformity became critical for individual survival. Individuals, therefore, benefited from developing reputations for norm compliance (by acting groupishly). Accordingly, we interpret groupishness as an evolved signal of willingness to follow norms, at a time when failure to signal such conformity would risk punishment, including ostracism and execution. This process is also expected to have been accompanied by strong selection on social–moral emotions, including embarrassment, guilt, and shame. Such emotions provided a within-individual system of self-monitoring that facilitated recognising, internalising, and redressing failures to act groupishly.
This review is not claiming that humans are unaggressive, or that humans only engage in coalitionary proactive aggression. Both reactive and proactive aggression are far too frequent and variable to permit such statements. For instance, bullying is a recurrent problem in various settings, infanticide remains prevalent, and female abuse occurs globally. Rather, our claim is that rates of most reactive aggression and some kinds of proactive aggression are much lower in humans than amongst chimpanzees.
These comparisons illustrate how socio-ecological conditions drive variations in species-typical aggression profiles, whilst also permitting acknowledgement of the continuing importance of restraining aggression in human societies.
