
The PLOS article by Marshall, Ruesseler, Hunt, O’Reilly “representation of priors and decisions in the human parietal cortex” discusses how both humans and animals actively sample the environment using their sensory organs, far from being passive recipients of sensory information. In rodents, active sampling processes include whisking and sniffing; in primates, the most important and best-studied process is the control of saccadic eye movements.
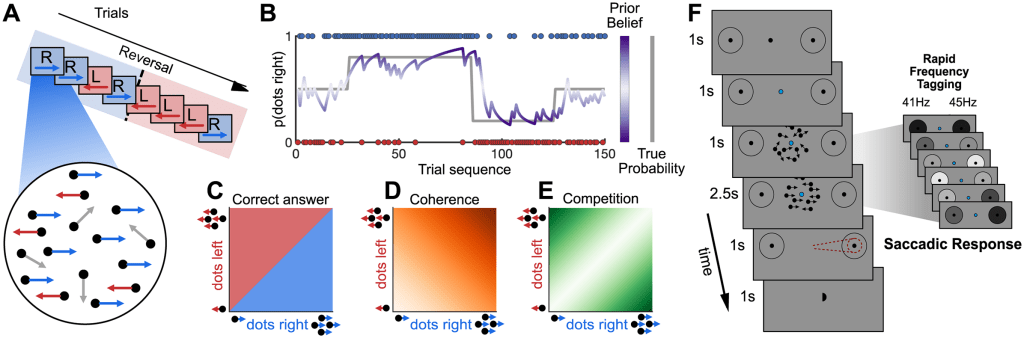
(A) Trial sequences were presented where certain motion directions were more common, with unpredictable “reversals” (top), and stimuli within a trial varied along 2 orthogonal dimensions; number of dots moving left and right, with all other dots moving randomly (bottom).
(B) The cross-trial prior probability p(correct direction = right) varied across the experiment, with pseudo-random, unsignalled “blocks” of trials in which the dominant direction was rightwards 20%, 50%, or 80% or the time (grey line). Bayesian learning models were used to estimate direction and strength of beliefs and observer should have about the current trial based on previous trials (purple line).
(C) On a given trial, the correct response is given by the dominant motion direction.
(D, E) Additionally, 2-d stimulus space can also be parameterized as varying along 2 dimensions: total coherence (middle panel), or the total percentage of coherent motion to the left or right, and competition (right panel), the unsigned difference between proportion of coherent dots moving left and right.
(F) Structure of a single trial: A get-ready cue indicated dots were about to appear. All trials began with 1 second of random motion, which temporally separated stimulus onset from evidence onset, meaning we were able to distinguish visual evoked neural responses from evidence accumulation processes. This also provided a temporally extended foreperiod in which neural activity reflecting prior beliefs could be measured. Random motion was followed by a 2.5-second evidence accumulation period, in which coherent motion was present. When the dots disappeared, participants had a 1-second response interval to make a saccade in the direction of perceived dominant motion. Participants were given unambiguous feedback on the correct answer (small centrally presented hemisphere on the correct side), so they could learn the cross-trial prior independently of the quality of evidence on each trial. During the 1-second foreperiod and 2.5-second period of evidence accumulation, the 2 potential saccadic targets were “tagged” with high frequency flicker to selectively entrain neural oscillations.
As an observer views a visual scene, several saccadic eye movements per second are generated in order to direct the eye’s small focal window to points of potential interest. However, each sample does not stand alone—instead, information from multiple fixations is integrated to construct a “model” of the full visual field, and this model, in turn, acts as a “prior,” influencing the selection of targets for future saccades. Therefore, the process of active sampling may be viewed as an interplay between 2 concurrent processes with distinct characteristics:
- Firstly, the brain must generate each individual sampling action. Since only one saccade is made at once, each individual saccade must be selected by a process of competition between representations of alternative possible saccadic targets; the process is by necessity winner-take-all in that the eyes can only fixate one location at a time and must operate on a fast timescale, driven by dynamics that ensure a new saccade is made every few hundred milliseconds.
- Secondly, the brain must integrate the limited information gained from many individual saccades—not only to construct the visual scene but relatedly to inform the selection of targets for future saccades. Behavioural modelling suggests that the likely information value of future saccades is represented in a salience map that can be regarded as a Bayesian prior distribution over potential saccadic targets. Far from having winner-take-all dynamics, this salience map must capture the relative or probabilistic distribution over all possible saccades. It therefore must integrate information across multiple previous saccades.
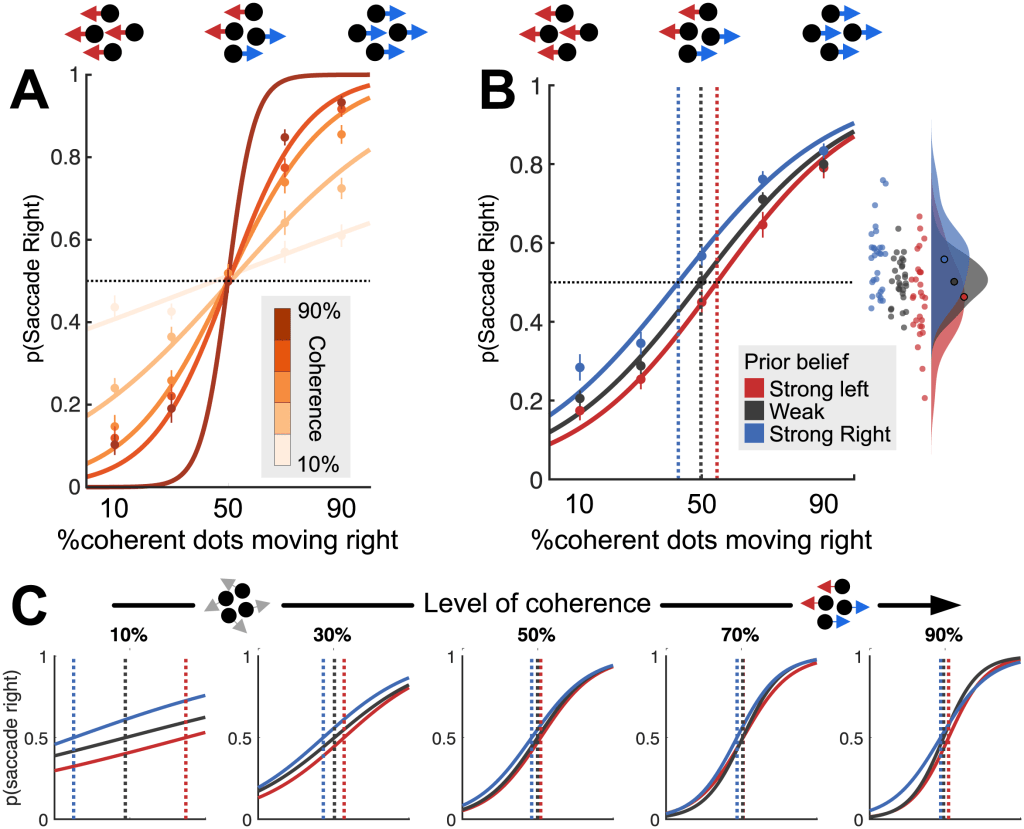
(A) Increasing overall coherence (darker lines) produces a parametric performance improvement (steeper logistic curve). Dots indicate mean observed data; lines indicate logistic fits.
(B) Prior belief influences choice behaviour on current trial, leading to a shift in the point of subjective equality (dashed vertical lines). Inset: Raincloud plot showing values of p(Saccade right) for individual participants on trials where 50% of dots moved right.
(C) Prior belief shifts point of subjective equality (dashed lines) most strongly when the least information is available on the current trial (coherence is lowest).
Our results provide evidence for a specific mechanism by which parietal cortex has been proposed to represent prior beliefs over longer timescales, namely, selective modulation of the sensory gain between visual and parietal cortices. Notably, and contradicting our expectations, we did not detect effects of the prior on the mean field, although this does not rule out the possibility that such an effect occurred.
Furthermore, we cannot rule out that active changes such as in baseline firing rate also occur, and, indeed, these have previously been observed; as bandpass filtering is a necessary step in the MEG processing pipeline, such tonic effects would be attenuated in our signal.
However, theory suggests that, when representations are sustained over long inactive periods or delays may be more efficiently represented by non-spiking mechanisms provide particularly energy-efficient ways to implement this. Physiologically, gain modulations can arise from a wide range of mechanisms pertaining to synaptic dynamics, neuromodulatory factors, and the timing and position of inputs. One possible mechanism is short-term synaptic plasticity on the timescale of seconds. This is reminiscent of the proposal that short-term synaptic plasticity may act as an activity-silent store for working memories; the activity pattern associated with working memory is later reactivated as a bump attractor.
Indeed, a prior over saccadic targets could be viewed, in terms of timescale, as a form of noncognitive working memory.
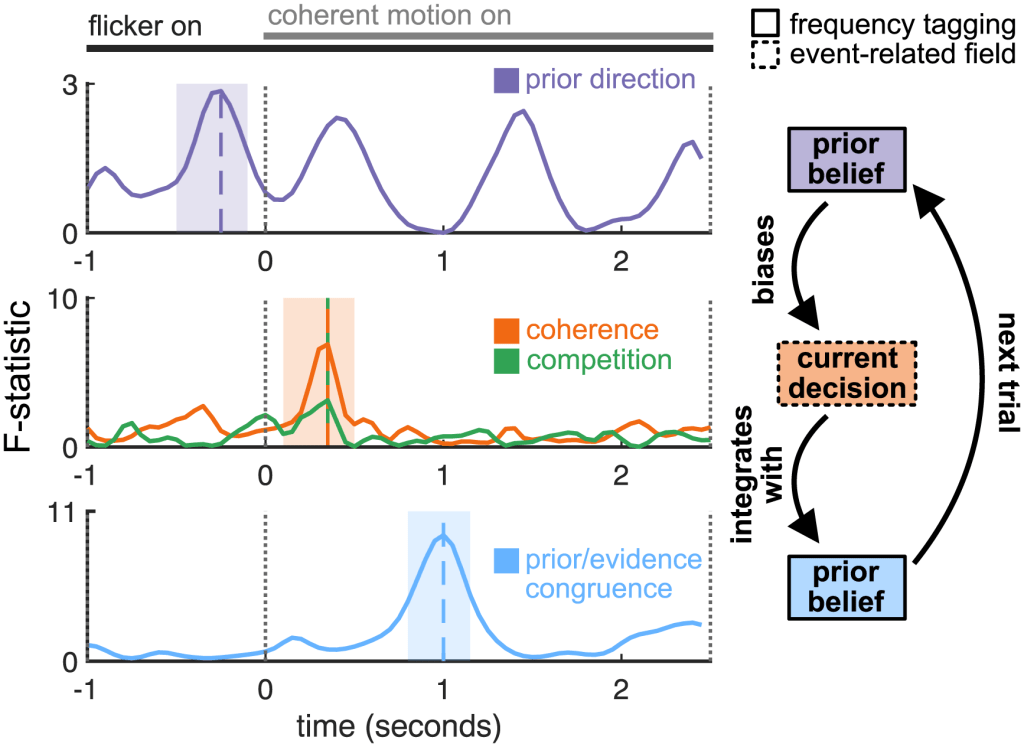
Temporal cascade of computations. Time courses of significant statistical effects; repeated measures ANOVAs comparing contralateral, weak, and ipsilateral prior belief (top row), all levels of stimulus coherence (middle row, orange) and stimulus competition (middle row, green) and comparing “evidence (dots) congruent with belief” trials with “evidence (dots) incongruent with belief” trials (bottom row). Bottom panel shows the time point-wise repeated measures ANOVA comparing dot-prior-congruent with dot-prior-incongruent trials (even though this is a paired comparison, an F- rather than t-statistic is shown to facilitate visual comparison with the upper and middle panels.
